
СИБИРСКИЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ
ИНСТИТУТ ЕСТЕСТВЕННЫХ И ГУМАНИТАРНЫХ НАУККАФЕДРА МИКРОБИОЛИГИИ
Послепожарная динамика микробных комплексов почв смешанных лиственничников Нижнего Приангарья
Выполнил: Филиппов А. М., студент 4 курса, гр. БФ05-31С
Научный руководитель: Богородская А.В., к.б.н.
Красноярск, 2009
Оглавление
Введение 3
Глава 1. Обзор литературы 5
1.1 Классификация лесных пожаров и их экологическая роль 5
1.2 Влияние пожаров на почвенный компонент лесного биогеоценоза 7
1.3 Влияние пожаров на почвенные микробоценозы 8
Глава 2. Объекты и методы исследования 12
Глава 3. Послепожарная динамика структуры и численности ЭКТГМ в почвах смешанных лиственничников 15
Глава 4. Послепожарная динамика содержания микробной биомассы и интенсивности базального дыхания почв смешанных лиственничников 19
Выводы 24
Литература 25
Введение
Почвенные микроорганизмы являются ключевым компонентом в процессах разложения растительных остатков и, следовательно, играют важную роль в питательных циклах. Полифункиональность и высокая чувствительность микроорганизмов к стрессам, позволяют использовать характеристики микробных комплексов почв для целей прогнозирования состояния лесных биогеоценозов после различных нарушений (Звягинцев, 1999).
Лесные пожары – основной и естественный фактор динамики и формирования лесных биогеоценозов (Валендик и др., 2000). В Сибири, где сосредоточена значительная часть бореальных лесов, ежегодно до 12-15 млн. га лесов повреждаются пожарами разной интенсивности (Conard, Ivanova, 1997).
Пожары оказывают влияние на все компоненты биогеоценоза. Почва, как неотъемлемая составная часть экосистемы, также подвергается сложному и разностороннему пирогенному воздействию, приводящему к изменению ее гидротермических и трофических условий (Попова, 1997; Краснощеков, 2004), а, следовательно, и биологических свойств (Сорокин, 1983; Pietikainen, Fritze, 1995; Nearly et. al., 1999; Wuthrich et. al., 2002; Certini, 2005). Изменения в почвах, происходящие после пожаров в значительной степени зависят от интенсивности пожара (Certini, 2005). Однако существует мало исследований, касающихся влиянию пожаров разной интенсивности на почвенные микробоценозы (Цветков и др., 2001; Богородская и др., 2006).
Основное количество пожаров по числу и площади приходится на южнотаежные леса Средней Сибири (Иванова, 2005). Одними из наиболее горимых являются леса Нижнего Приангарья, где до 22,6% лесопокрытой площади приходится на насаждения с участием лиственницы (Попов, 1982). Однако, влияние пожаров разной интенсивности на микробоценозы почв лиственничников южной тайги остается неизученной областью.
Цель исследования заключалась в изучении послепожарной динамики характеристик микробных комплексов почв смешанных лиственничников южной тайги Средней Сибири.
В задачи исследования входило:
1.исследовать структуру и численность эколого-трофических групп микроорганизмов (ЭКТГМ) после пожаров разной интенсивности в смешанном лиственничнике;
2. проследить послепожарную динамику восстановления функциональной активности микробоценозов почв изучаемых лиственничников.
Глава 1. Обзор литературы
1.1 Классификация лесных пожаров и их экологическая роль
Лесной пожар – это горение, стихийно распространяющееся по лесной территории (Курбатский, 1962; 1970).
В зависимости от сгорающих материалов различают три основных вида лесных пожаров: низовые, верховые и почвенные или подземные (Курбатский, 1970). Наиболее распространенные низовые пожары. При низовых пожарах гибнут нижние наземные ярусы фитоценоза, живой напочвенный покров, подрост и подлесок. Различают беглые и устойчивые низовые пожары. К беглым относят пожары с быстро продвигающейся кромкой (скорость распространения более 0,5м/мин), когда сгорает лишь напочвенный покров, опад, подрост и хвойный подлесок. При устойчивом пожаре на участке длительное время продолжают гореть подстилка, валежник, гнилые пни. Участок при этом сильно дымит.
К верховым относятся пожары, при которых наряду с нижними ярусами растительности сгорает полог древостоя. Эти пожары возникают из низовых, как дальнейшая стадия его развития, причем низовой огонь является составной частью верхового пожара. При верховом пожаре древостой гибнет полностью.
Среди почвенных пожаров различают два вида: подстилочные и торфяные. Это деление обосновано особенностями горючих материалов: в одном случае горит подстилка, а в другом – слой торфа, причем его горение беспламенное.
Пожары неодинаковы по своей повторяемости интенсивности и развитию. Эти характеристики в первую очередь определяются климатическими условиями, аккумуляцией горючих материалов и их способностью к загоранию, почвами и особенностями рельефа местности.
Интенсивность и сила пожара описывают его поведение и последствия. Интенсивность – это физическая характеристика поведения пожара, определяемая как количество тепла, выделяющегося с единицы длины кромки фронта в единицу времени (Курбатский, 1972).
Сила пожара, в широком смысле, определяется как степень изменения экосистемы, под воздействием пожара. Сила пожара может быть определена через количество древесного отпада (Agee, 1993), теплоту, проникающую в почву при пожаре (Lea and Morgan, 1993), количество поглощенной огнем биомассы (Lenihan et. al., 1998) или через сочетание перечисленных последствий (Turner et. al., 1994).
Отклик экосистем на воздействие пожаров проявляется по-разному и зависит от вида и интенсивности пожара, климатических особенностей региона, строения древостоя, структуры нижних ярусов растительности и других факторов.
Пожары всегда были естественными и крайне важным фактором окружающей среды. Как экологический процесс, пожары оказывают влияние на динамику растительности и ее продуктивность (Фуряев и др., 2001; Валендик и др, 2000; Фуряев и др., 2004;), животные популяции (Ahlgren, 1974), минеральный и углеродный циклы (Sofronov, Volokitina, 2000), почвенные биологические процессы (Ahlgren, 1974). Результатом подобных воздействий стало преобразование бореальных лесов из стока углерода в его источник (Apps, 2000).
Установлено, что в результате пожаров любой интенсивности в каждом природно-территориальном комплексе (ПТК) прослеживается следующая цепь взаимосвязанных и взаимообусловленных явлений. Полное уничтожение огнем подстилки, живого напочвенного покрова, подлеска, подроста и древесного яруса обуславливает резкое повышение освещенности поверхности почвы, увеличивает проникновение осадков на ее поверхность и их интенсивность, изменяет температурный режим почвы и влажность приземного слоя воздуха (Фуряев, 1996), интенсифицирует процесс инфильтрации солей и мелких частиц, изменяет направленность почвообразовательного процесса, перераспределяет поверхностный и внутрипочвенный стоки, изменяет уровень грунтовых вод и их трофность, перераспределяет элементы минерального питания между различными ПТК (Сапожников, 1979).
Таким образом, средопреобразующая роль пожаров в лесных биогеоценозах, обуславливающая изменение экологических режимов и особенности послепожарного лесообразовательного процесса, неоднозначна и зависит как от характеристик пожара и их повторяемости, так и от исходного типа леса.
1.2 Влияние пожаров на почвенный компонент лесного биогеоценоза
Почва, как главный компонент биогеоценоза, наиболее чувствителен к воздействию пожаров. Помимо прямого (пиролиз) пожары оказывают огромное косвенное влияние на лесные биогеоценозы. Они коренным образом меняют эдафические условия, а значит, микробиологические и биохимические процессы в почвах (Сапожников и др., 1993).
Воздействие пожаров на почву идет несколькими путями. Это – непосредственное воздействие высоких температур на твердую фазу почв, единовременное поступление на поверхность почвы значительного количества золы, образовавшейся при минерализации подстилки и других горючих материалов, изменение структуры и качества органического вещества и смена одних растительных сообществ другими (Попова, 1997).
Можно отметить, что в результате пожаров существенно изменяются физико-химические свойства, механический состав, водно-воздушный и гидротермический режим почв, что оказывает непосредственное влияние на биологические свойства почв (Краснощеков, 2004; Certini, 2005).
Большую роль в развитии низовых пожаров, по статистке составляющих до 90% от общего числа пожаров, и их влиянии на почву имеет лесная подстилка, которая является основным видом горючего материала и максимально приближена к минеральным горизонтам. Если подстилка имеет большую мощность, что часто наблюдается в хвойных лесах, то при пожаре сгорает лишь ее верхний сухой слой, а нижний, как более влажный, – сохраняется. Поэтому при такой ситуации прямой эффект воздействия пожара на почву незначителен. Полностью же подстилка может быть уничтожена огнем только при сильной засухе или при многократном повторении пожаров в течение нескольких лет. При сильном воздействии огня и полном сгорании подстилки происходит также трансформация верхних органно-минеральных и минеральных горизонтов почв, которые обнажаются, что способствует развитию эрозии (Валендик и др., 2000).
1.3 Влияние пожаров на почвенные микробоценозы
Почвенные микробоценозы представлены многочисленными группами микроорганизмов, связанных сложными трофическими взаимоотношениями. Изучение структуры и численности эколого-трофических групп микроорганизмов, их разнообразия, биохимической активности почвы позволяет судить о степени выгорания органогенного горизонта, послепожарных изменений лесорастительных свойств почв, изменений трофических условий почвенного ценоза.
Одним из основных почвенных компонентов, реагирующих на пирогенное воздействие, являются микробоценозы, что проявляется в изменении структуры и функциональной активности (Сорокин, 1983). Так, на гарях 4- и 16-летней давности абсолютное содержание всех эколого-трофических групп микроорганизмов в 1,5-3,0 раза выше, чем в контроле и в 3-7 раз, чем на однолетней гари (Цветков и др., 2001).
Огонь может влиять на почвенные микробные популяции и видовой состав в зависимости от интенсивности пожара, максимальных температур, от типа почвы и увлажнения, продолжительности огневого воздействия и глубины прогорания, от до и послепожарного состава растительного яруса и климатических условий (Hungerford et. al., 1995; Certini, 2005). Количество горючих материалов, месторасположение участка, его возвышенность и до и послепожарные погодные условия также важны, так как они могут влиять на поведение огня и дальнейшее восстановление. Низкоинтенсивные, с быстропродвигающейся кромкой огня пожары не оказывают сильного влияния на почвенные микробные популяции. Высокоинтенсивные пожары могут стать причиной значительных изменений в микробных популяций почв (Nearly et. al., 1999).
Влияние прогревания среды на микроорганизмы многосторонне, особенно в такой гетерогенной среде как почва. Обычно, при прогревании почвы количество микроорганизмов в ней в первое время уменьшается. Несмотря на то, что при низовых лесных пожарах температура поверхности подстилки колеблется в пределах от 930 до 7200С, температура верхнего трех-четырех сантиметрового слоя почвы не превышает 50-800С. Незначительные различия в популяции почвенных микроорганизмов до и сразу после прогревания могут быть объяснены относительно низкими температурами почвы (Ahlgren, 1974). Температура свыше 1270С стерилизует почву, а воздействие 700С убивает неспорообразующие грибы, и некоторые бактерии, но практически не влияет на спорообразующие
Пожары средней и низкой интенсивности убивают неспорообразующие грибы, простейших, актиномицетов и некоторые бактерии, но практически не влияют на спорообразующие бактерии и грибы (Bacillus spp., Clostridium spp.). Несмотря на то, что общее количество бактерий в почве сразу после пожаров средней и низкой интенсивности обычно уменьшается, тем не менее, через некоторое время их количество увеличивается благодаря быстрому росту. Очевидно, это связано с внезапным увеличением количества минеральных питательных веществ, которые постепенно вымываются в более глубокие слои, увеличения рН почвы и прочих химических изменений, связанных с горением. Повторный засев может произойти очень быстро после пожара за счет принесенных ветром спор и прочих остатков и за счет вторжения из подпочвенных слоев. Большое значение при этом имеет влага: увлажнение почвы и выпадение дождей после пожара оказывает благоприятное действие на повторный засев и увеличение популяций микроорганизмов (Vazqeuz et. al., 1993).
При пожарах высокой интенсивности наблюдается снижение микробиологической деятельности, а при средней и низкой интенсивности огня жизнедеятельность микроорганизмов усиливается (Гуняженко, 1970; Попова, 1978). После обжига отмечается смещение зоны действия микроорганизмов в нижележащие слои почвы. По истечению двух-трех лет после пожаров низкой и средней интенсивности разница между обожженной и необожженной почвой сглаживается (Сорокин, 1983).
Прогрев почвы по глубине является основным фактором воздействия огня на свойства почв и микрофлору. Степень нагревания почвы часто определяет послепожарную растительность, влияет на водопроницаемость почвы и образование эрозии. Температура на поверхности почвы при пожаре может превышать 900оС, доходить до 1500оС при высокоинтенсивных пожарах, а температура в 200-300оС является обычной при горении (Davis, 1959). Основная доля тепла при горении уносится нагретыми газами в атмосферу, а рассеивание энергии теплопроводностью составляет небольшую долю от всего расхода тепла. По некоторым оценкам рассеивание тепла низовых пожаров в окружающую среду в среднем составляет: излучение в стороны – 18-20%, по некоторым данным – 25%, конвекцией и излучением вверх – 70-80%, теплопроводностью в почву – 3-5% (Humphreya, Craig, 1981).
Следовательно, действие пирогенного фактора распространяется до глубины 10 см, где микроорганизмы гибнут при высокой интенсивности огня (Grasso et. al., 1996; Neary et. al., 1999; Certini, 2005).
Дополнительным буфером, предохраняющим почву от действия высоких температур, служит лесная подстилка, имея высокое влагосодержание, она плохо горит и снижает термическое воздействие на почву (Курбатский, 1962; Campbell et. al., 1995). При пожаре сгорает только часть подстилки, почва и нижняя часть подстилки при этом, как правило, остаются влажными. В такой ситуации прямой эффект на почву незначителен (Grasso et. al., 1996).
Таким образом, резкое подавление микробиологической деятельности в почве наблюдается лишь при очень сильных пожарах, когда подстилка выгорает полностью (Попова, 1986). Но и в этих случаях снижение активности микроорганизмов бывает временным.
Влияние пожара на микробные комплексы почв может быть опосредованным. На скорость восстановления микробоценозов почв после пожаров влияет удаление или сокращение источников органики, изменение качественного состава органических субстратов, физико-химических и гидротермических свойств почв (Fritze, Pietikainen, 1993; Pietikainen, Fritze, 1993, 1995; Nearly et. al., 1999; Краснощеков и др., 2004; Certini, 2005).
Глава 2. Объекты и методы исследованияИсследования микробоценозов почв проводились в подзоне южной тайги (Нижнее Приангарье) на левом берегу р. Ангара в 30 км вверх по течению (58о35' с.ш. и 98о55' в.д.). Территория исследования по геоморфологическому районированию относится к Приангарскому понижению Енисейского кряжа. Рельеф представлен плоскими и полого-холмистыми плато с реликтами неогеновой аллювиальной равнины, характеризуется значительной расчлененностью с колебаниями отметок от 100 до 450 м (Козловская, 1971). Климат района резко континентальный. Среднегодовая температура воздуха колеблется в пределах от –2.0оС до –2.4оС. Безморозный период длится в среднем 103 дня. Годовая сумма осадков составляет 320-380 мм осадков (Галахов, 1964). В почвенном покрове господствуют дерново-глубокоподзолистые почвы, своеобразие которых заключается в большой мощности элювиальной толщи и высокой степени оподзоленности.
Исследования послепожарной динамики микробных комплексов почв проводились на 2 экспериментальных участках (площадью 1 га), пройденных пожарами высокой интенсивности в 2006 году (№2) и низкой интенсивности в 2007 году (№3) и на контроле к ним.
Тип леса - лиственничники мелкотравно-зеленомошные. Состав древостоя имеет сложную структуру 3Л2С1К2Е1П1Б, Ос. Возраст древостоев 140 лет, средняя высота 27 м. Сомкнутость крон II яруса - 0.7-0.9, возраст 40 лет, средняя высота 16 м. Верхний ярус полидоминантный, с явным доминированием Larix sibirica Ledeb. и Pinus sylvestris L. В составе второго яруса преобладают Abies sibirica Ledeb., Picea obovata Ledeb., Pinus sibirica (Du) Tour. Подрост темнохвойный, крупномерный более 3 тыс. шт./га. Подлесок как ярус не выражен, единично представлены Salix caprea L., Rosa acicularis Lindl., Spiraea media Franz Schmidt, Lonicera tatarica L., Sorbus sibirica Hedl., Juniperus communis Burgsd. Общее проективное травяно-кустарничкового яруса составляет от 10 до 50%, где доминируют Linnaea borealis L., Pyrola rotundifolia L., Viola uniflora L., Anemone altaica Fischer ex C. A. Meyer, Lathyrus humilis (Ser.) Spreng., Rubus saxatilis L., Carex macroura Meinsh., Vaccinium vites-idea L. Общее проективное покрытие мхов составляет 40—80%, где доминируют Pleurozium schreberi (Brid.) Mitt, Hylocomium splendens (Hedw.) Schimp. B.S.G. в примеси Dicranum polysetum Michx. Лишайниковый покров не развит.
Почва характеризуется как бурозем темный выщелоченный маломощный глинистый на элювиально-делювиальной красноцветной мергелистой глине (Классификация…., 2004).
Образцы почвы для микробиологических анализов были взяты в пяти точках на каждом участке до и через сутки после выжигания, а также в течение последующих двух лет в середине вегетационного периода. Параллельно были отобраны образцы на контрольном участке, не пройденном огнем. Точки отбора были постоянными. Отбор образцов проводили по следующим слоям: подстилка (О), 0-5 - маломощный гумусовый горизонт коричневато-черного цвета, 5-15 см - переходный горизонт.
Характеристику микробных сообществ почв проводили на основе изучения структуры и численности эколого-трофических групп микроорганизмов (ЭКТГМ) (Методы…, 1991). Численность аммонификаторов изучали на мясо-пептонном агаре (МПА), прототрофов, использующих минеральные формы азота – на крахмало-аммиачном агаре (КАА), олиготорофов – на почвенном агаре (ПА) и грибы – на сусло-агаре с молочной кислотой (СА).
Содержание микробной биомассы (Смик) определяли методом субстрат-индуцированного дыхания (СИД) (Anderson, Domsh, 1979; Ананьева, 2003). В пенициллиновые флаконы объемом 14.5 мл помещали 2 г свежей почвы с тщательно отобранными корнями или 1 г подстилки, добавляли 0.2 мл глюкозо-минеральной смеси (ГМС мг/мл: глюкоза - 200; К2НРО4 - 20; (NH4)2SO4 - 20), увлажняли при необходимости до 60% от полной влагоемкости, герметично закрывали пробками и инкубировали при 22С в течение 4-6 часов подстилку и 8-12 часов почву. Длительность инкубирования определялась согласно продолжительности лаг-периода роста микроорганизмов (отдельно для микробоценозов почвы и подстилки) на ГМС.
Образование СО2 почвой оценивали с использованием газового хроматографа (ЛХМ80 модификация «Хром 4»). Смик определяли путем пересчета скорости СИД по формуле (Anderson, Domsh, 1979):
Смик (мкг г-1 почвы) = (мкл СО2 г-1 почвы час-1)Ч40.04+0.37.
Базальное (фоновое) дыхание (БД) почвы измеряли по скорости выделения СО2 почвой за 120 ч ее инкубации при 25оС. Определение СО2 проводили хроматографически, как описано для определения СИД, только вместо внесения раствора ГМС, вносили воду. Скорость базального дыхания выражали в мкг СО2-С / г почвы в час.
Статистическую обработку данных выполняли с использованием компьютерной программы Microsoft Excel 2002.
Глава 3. Послепожарная динамика структуры и численности ЭКТГМ в почвах смешанных лиственничников
Изучение структуры и численности основных групп микроорганизмов бурозема темного выщелоченного показало, что максимальная их численность расположена в подстилке и верхней части темногумусового горизонта и постепенно убывает вниз по профилю, что характерно для автоморфных почв (рис. 1). Преобладание прототрофов над аммонификаторами указывает на высокую интенсивность процессов микробиологической минерализации органических веществ. В то же время, численность олиготрофов, извлекающих мономерные соединения при низкой их концентрации в среде, достаточно высока, но чаще всего не превышает количество аммонификаторов, что свидетельствует об оптимальном количестве легкоминерализуемого органического вещества для поддержания пула гидролитической и копиотрофной группировок в микробном комплексе (рис. 1). Следует отметить невысокую численность микромицетов в составе микробных комплексов почв (2.2-0.5 млн. КОЕ/1г) и высокое процентное содержание актиномицетов (до 10-15%), выросших на крахмало-аммиачном агаре. В целом отмечена достаточно высокая вариабельность численности микроорганизмов в подстилке и верхней части темногумусового горизонта лиственничника, что объясняется мозаичностью напочвенного покрова и влиянием нано- и микрорельефа.
Выявлено, что действие пожара высокой интенсивности на микробоценозы почвы смешанного лиственничника неоднозначно в пределах одного участка: изменение структуры и численности, а также функциональной активности микробоценоза зависит от степени прогорания подстилки с напочвенным покровом (НП) во время пожара, которая сильно варьирует и определяется неравномерным распределением горючих материалов, мозаичным характером НП и влажностью почвы, связанными с микрорельефом. Сильному прогоранию соответствовали зоны со значительным сгоранием подстилки и НП (4-5 см), среднему – 1,5-3 см, слабому – менее 1 см. Зоны слабого прогорания незначительны на участке 2 и приурочены к микропонижениям с более высоким увлажнением подстилки, которая слабо прогорела.
Самые значительные изменения численности микрофлоры после пожара происходят в подстилке, которая подвергается воздействию высоких температур во время пожара (около 1000оС). Максимальное снижение численности микроорганизмов изучаемых эколого-трофических групп (на 50-80%) отмечено в подстилке и верхней части темногумусового горизонта (0-5 см) в зонах с сильным прогоранием (рис. 1). В зонах со слабым прогоранием численность микрофлоры изменяется незначительно по сравнению с допожарным уровнем. Следует обратить внимание на возросшее здесь в 10 раз количество грибных зачатков (в основном за счет Penicillium), что, вероятно, связано с созданием благоприятных гидротермических условий при тепловой мелиорации достаточно влажной подстилки в совокупности с обогащением почвы легкогидролизуемым органическим веществом и зольными элементами (Попова, 1997; Fritze, Pietikainen, 1993; Certini, 2005).
В верхней части темногумусового горизонта (0-5 см) после пожара во всех зонах количество аммонификаторов доминирует над численностью прототрофов, либо равно их количеству, что свидетельствует о снижении темпов микробиологической минерализации остаточного органического вещества. В нижней части того же горизонта (5-15 см) в зонах с сильным прогоранием подстилки и НП количество микроорганизмов несколько повышается, а в зонах со средним и слабым прогоранием не наблюдается значительных изменений структуры и численности ЭКТГМ, поскольку на данной глубине температура во время пожара не поднимается до критических для микрофлоры значений (Богородская, 2006).

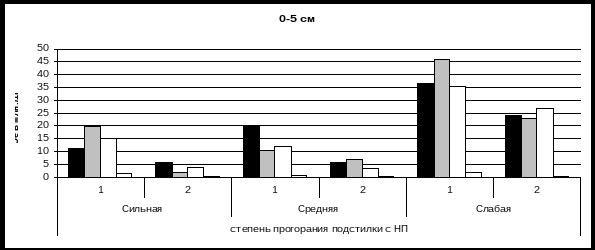
 Рис. 1. Послепожарная динамика структуры и численности ЭКТГМ в почве лиственничника после пожара высокой интенсивности (уч. 2) 1 – до пожара; 2 – через сутки после пожара.
Рис. 1. Послепожарная динамика структуры и численности ЭКТГМ в почве лиственничника после пожара высокой интенсивности (уч. 2) 1 – до пожара; 2 – через сутки после пожара.
Сразу после низкоинтенсивного пожара (участок №3) в подстилке количество аммонификаторов и прототрофов несколько повышается, тогда как количество грибных зачатков уменьшается в 2 раза (рис. 2). В верхнем 5 см слое темногумусового горизонта отмечено незначительное снижение аммонификаторов и почти трехкратное уменьшение численности грибной микрофлоры. Ряд авторов указывает на большую по сравнению с бактериальной флорой чувствительность микромицетов к воздействию высоких температур (Dunn et. al., 1985; Diaz-Ravina et. al., 1996; Fritze, Pietikainen, 1993).
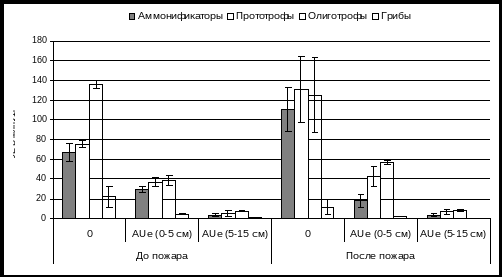
Рис. 2. Численность микроорганизмов в буроземе темном оподзоленном после низкоинтенсивного пожара (уч.№3)
Глава 4. Послепожарная динамика содержания микробной биомассы и интенсивности базального дыхания почв смешанных лиственничников
К основным характеристикам функционирования микробных комплексов относят величину микробной биомассы и показатель активности функционирования микробоценозов – выделение углекислоты. Отношение этих показателей позволяет давать дополнительную оценку состояния микробных комплексов после различных нарушений (Ананьева, 2003).
Фундаментальное значение живой микробной биомассы в биотическом круговороте лесных экосистем связано с биологическим окислением (разложением) полимеров растительного происхождения. Использование гетеротрофными микроорганизмами (бактерии, грибы, актиномицеты) в качестве источников углерода и энергии самых разнообразных субстратов, обеспечивает до 80-90% минерализации общей первичной продукции наземных экосистем (Стейниер и др., 1979).
Содержание общей микробной биомассы в подстилках и почвах лесных экосистем – показатель не стабильный и, зависящий от многих факторов. В первую очередь на значение этого показателя оказывают влияние гидротермические условия и наличие в почве доступных органических веществ (Ананьева, 2003).
Скорость продуцирования углекислоты необогащенной почвой – базальное дыхание (БД) или микробное дыхание, может служить показателем изменений состояния микробоценозов почв после различных нарушений (Ананьева, 2003).
Таким образом, к основным функциональным характеристикам микробных комплексов можно отнести величину микробной биомассы и показатель активности функционирования микробоценозов – выделение углекислоты. Взаимосвязь этих показателей позволяет давать оценку состояния микробных комплексов после различных нарушений (Anderson. Domsch, 1993; Ананьева, 2003).
Для оценки функциональной активности микробного комплекса почвы после пожара использовали содержание микробной биомассы (Смик) и интенсивность базального дыхания (БД) почвы.
Выявлено, что действие пожара высокой интенсивности на функциональную активность микробоценозов почвы смешанного лиственничника неоднозначно в пределах одного участка и также как и численность зависит от степени прогорания подстилки с напочвенным покровом (НП). Установлено, что после высокоинтенсивного пожара (участок 2) изменение вышеназванных величин на участке варьирует и зависит от степени прогорания подстилки (рис. 3). Максимальное снижение углерода микробной биомассы наблюдается в подстилке с сильным ее прогоранием (на 85%) и минимальное – в зонах со слабым прогоранием подстилки (в 1.5 раза). В зоне со средним прогоранием подстилки величина Смик практически не менялась.
В гумусо-аккумулятивном слое почвы (0-5 см) содержание Смик уменьшается в 2 раза в зонах с сильным прогоранием, в то время как в зонах со средней и низкой степенью прогорания подстилки - снижается незначительно. В нижележащем слое почвы (5-15 см) послепожарные изменения Смик не так наглядны.
Послепожарное снижение микробной биомассы в работах Pietikainen, Fritze (1993, 1995), Fritze et. al. (1998) и Pietikainen et. al. (2000) объясняется как снижением уровня влажности почвы, так и изменением структуры почвенного органического вещества, что также оказывает влияние на развитие микробной биомассы в почвах после пожаров.
Интенсивность БД в подстилке и гумусо-аккумулятивном слое почвы практически не меняется в зонах с сильным прогоранием подстилки, тогда как в зонах со средним и слабым прогоранием либо не меняется, либо увеличивается в 1.5 раза.
Через год после высокоинтенсивного пожара отмечено обильное зарастание гари кипрейно-злаковой группировкой. В подстилке и гумусо-аккумулятивном слое содержание микробной биомассы снижено в 1,5-2 раза только в зоне с сильным прогоранием подстилки, тогда как в зоне со средним и слабым прогоранием содержание Смик сравнимо с контролем. Интенсивность микробного дыхания в подстилке и верхнем слое почвы (0-5 см) сгоревшего лиственничника ниже примерно в 1,5 раза во всех зонах. В нижележащем слое 5-15 см параметры Смик и БД ниже контроля почти в 2 раза в зонах с сильным и средним прогоранием подстилки с НП.
Через два года после высокоинтенсивного пожара содержание Смик в подстилке и слое 0-5 см снижено почти в 2 раза только в зоне с сильным прогоранием, тогда как в зонах со средним и слабым прогоранием - выше контроля в 1,5-1,8 раз. Интенсивность микробного дыхания в подстилке и слое 0-5 см в зоне с сильным прогоранием ниже контроля в 2-3 раза, в зоне со слабым прогоранием интенсивность БД увеличивается в подстилке, а в зоне со средним прогоранием - сравнима с контролем.
Сразу после пожара низкой интенсивности (уч. № 3) содержание микробной биомассы и базального дыхания в подстилке практически не меняется, а в гумусо-аккумулятивном слое почвы 0-5 см содержание Смик повышается на 30%, а интенсивность БД увеличивается почти в 2 раза. В нижележащем слое почвы 5-15 см содержание Смик также несколько увеличивается, а дыхание микроорганизмов возрастает более чем в 2 раза. Послепожарная активизация функциональной активности микробоценозов связана с прогреванием почвы после низкоинтенсивного пожара. Так, А.В. Богородской (2006) было показано, что в момент горения на поверхности напочвенного покрова при высокоинтенсивном пожаре температура поднималась более 1000С, под подстилкой она кратковременно достигала 190С и не превышала 40С при низкоинтенсивном пожаре. В почве на глубине 5-10 см при высокоинтенсивном пожаре температура могла на несколько секунд подниматься до 100С, но в основном, не превышала 50С. В большинстве случаев она поднималась лишь на 2-3 С.
Увеличение интенсивности микробного дыхания почвы после пожаров можно объяснить обогащением почвы зольными элементами и продуктами пиролиза, богатыми азотом и доступной органикой (Kutiel et. al., 1989; Baath et. al., 1995; Diaz-Ravina et. al., 1996). Оставшиеся корни сгоревшей растительности также могут являться питательным субстратом для микрофлоры почв, обуславливая более высокое базальное дыхание (Wuthrich et. al., 2002).
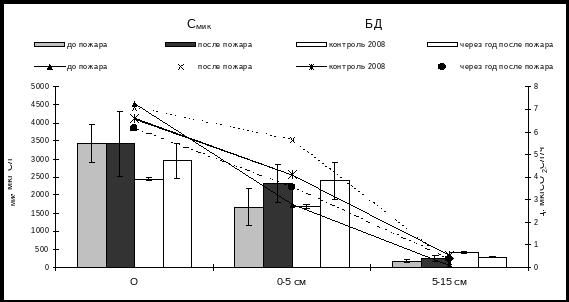 Рис. 4. Динамика микробной биомассы и базального дыхания в почве южнотаежного лиственничника после низкоинтенсивного пожара (уч. №3)
Рис. 4. Динамика микробной биомассы и базального дыхания в почве южнотаежного лиственничника после низкоинтенсивного пожара (уч. №3)
Через год после низкоинтенсивного пожара содержание углерода микробной биомассы в подстилке и слое 0-5 см несколько выше по сравнению с контролем, тогда как интенсивность микробного дыхания немного снижена (рис. 4).
Известно, что содержание Смик и интенсивность БД в значительной степени зависят от влажности и температуры почвы (Ананьева, 2003; Кожевин, 1989). Изменение величин Смик и БД в почвах изучаемых лиственничников существенно зависели от ее влажности: наиболее тесно с ней коррелировало содержание Смик (R2=0,65 при Р=0.05) и несколько слабее с влажностью связь интенсивности БД (R2=0.58 при Р=0.05).
Таким образом, изучение функциональных характеристик состояния микробоценозов почв сразу после пожаров показывают, что максимальные изменения активности функционирования микроорганизмов отмечаются после высокоинтенсивного пожара, а минимальные – после низкоинтенсивного.
Действие пожара высокой интенсивности на микробоценоз бурозема темного оподзоленного в смешанном лиственничнике Нижнего Приангарья неоднозначно в пределах одного участка: величина послепожарных изменений численности, биомассы и активности микроорганизмов зависит от степени прогорания подстилки и напочвенного покрова во время пожара, которая сильно варьирует и определяется неравномерным распределением горючих материалов, мозаичным характером произрастания напочвенного покрова и микрорельефом. Наибольшему воздействию пожара подвергались микробоценозы подстилки и верхнего 5 см слоя темногумусового горизонта в зонах с сильным прогоранием: отмечено уменьшение на 50-80% численности микроорганизмов изучаемых эколого-трофических групп, снижение на 85% углерода микробной биомассы в подстилке и в 2 раза в верхнем слое темногумусового горизонта. Интенсивность микробного дыхания сразу после пожара либо не менялась, либо незначительно увеличивалась.
В последующие послепожарные годы функциональная активность микробоценозов почвы смешанного лиственничника снижена только в зонах с сильным прогоранием подстилки с НП.
Пожары низкой интенсивности в смешанном лиственничнике в первый год приводят к активизации микробиологических процессов минерализации органического вещества, но уже через год функциональная активность бурозема темного оподзоленного сравнима с контролем.
ЛитератураAnderson J.P.E., Domsch K.H. A physiological method for the quantitative measurement of microbial biomass in soils // Soil Biol. and Biochem. 1978. V. 10. № 3. P. 314-322.
Baath E. Microbial community structure and pH response in relation to soil organic matter quality in wood-ash fertilized, clear-cut or burned coniferous forest soils / E. Baath, A. Frostegard, T. Pennanen, H. Fritze // Soil Biology and Biochemistry. - 1995. - 25. - P. 229-240.
Diaz-Ravina M. Bacterial activity in a forest soil heating and organic amendments measured by the thymidine and leucine incorporation techniques / M. Diaz-Ravina, A. Prieto, E. Baath // Soil Biology and Biochemistry. - 1996. - 28. - P. 419-426.
Dunn P. H. Soil moisture affects survival of microorganisms in heated chaparral soil / P. H. Dunn, S. C. Barro, M. Poth // Soil Biology and Biochemistry. - 1985. - 17. - Р. 143-148.
Kutiel P. Effects of simulated forest fire on the availability of N and P in Mediterranean soils / P. Kutiel, A. Shaviv // Plant and soil. - 1989. - 1. - P.57-63.
Vega G.A. Preliminary results of a study on short-term effects of prescribed fire in pine stands in NW Spain / G.A. Vega, S. Bara, M. Alonso, M.T. Fonturbel // Ecology Mediter. - 1987. - 13. - P. 177-188.
Wuthrich C. Soil respiration and soil microbial biomass after fire in a sweet chestnut forest in southern Switzerland / C. Wuthrich, D. Schaub, M. Weber, P. Marxer, M. Conedera // Catena. - 2002. - 48. - P. 201-215.
Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М.: Наука, 2003. 222 с.
Богородская А.В. Влияние пожаров на микробные комплексы почв сосновых лесов Средней Сибири. Автореф…. канд. биол. наук. Красноярск,: ИЛ СО РАН. 2006. 22 с.
Звягинцев Д.Г. Вертикально-ярусная организация микробных сообществ лесных биогеоценозов / Д.Г. Звягинцев, И.П. Бабьева, Т.Г. Добровольская, Г.М. Зенова, Л.В. Лысак, Т.Г. Мирчинк // Микробиология. - 1993. - Т.62. - Вып.1. - C. 256-278.
Иванова Г.А. Зонально-экологические особенности лесных пожаров в сосняках Средней Сибири: Автореф. дис. … докт. биол. Красноярск: ИЛ, 2005. 40 с.
Классификация и диагностика почв России /авторы и составители: Шишов Л.С., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. - Смоленск: Ойкумена, 2004. 342 с.
Попов Л.В. Южнотаежные леса Сибири. Иркутск: Изд-во Иркутского ун-та. 1982. 330 с.