
Оглавление:
1) Оглавление……………………………………………..02
2) Общие, разновидности………………………………..03
3) Система определения пола…………………………..03
4) Шмель: много шума ни из чего……………………..08
5) Укус шершня убил рыбака………………………….11
6) Приложение………………………………………12 - 17
Отряд перепончатокрылые (Hymenoptera) объединяют свыше 70 тыс. видов пчёл, шмелей, ос, наездников, муравьев некоторых других групп насекомых (рис. 1) . За исключением бескрылых форм, перепончатокрылые имеют две пары перепончатых, сравнительно небольших крыльев, из которых задние заметно меньше передних. На крыльях имеется небольшое число жилок, образующих крупные ячейки. У большинства видов крылья имеют радужный переливчатый своеобразный оттенок. Часто грудь соединяется с брюшком тонким стебельком. На конце брюшка самок обычно имеется яйцеклад или жало, являющееся видоизмененным яйцекладом. Ротовые органы грызущие или грызуще-сосущие.
Многие перепончатокрылые живут обществами, которые являются потомством одной матки. У многих общественных перепончатокрылых наблюдается полиморфизм. Их семьи состоят из одной или нескольких самок (маток), некоторого количества самцов и многих рабочих особей (бесплодных самок). Этим насекомым свойственна сложная нервная деятельность, в которой преобладают инстинктивные действия.
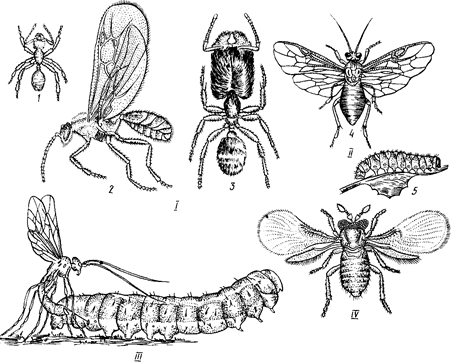
Рис. 1. Перепончатокрылые насекомые:
/ — муравьи; // — крыжовниковый пилильщик; /// — наездник парализует гусеницу бабочки перед откладкой на нее яиц; IV— яйцеед-трихограмма;
/ -- рабочий; 2 -- крылатый самец; 3 — солдат; 4 — взрослая самка; 5 — личинка
Насекомые, принадлежащие к группе, известной под названием перепончатокрылых (Hymenoptera), к которым относятся муравьи, пчелы и осы, обладают своеобразной системой определения пола. Термиты, не относящиеся к перепончатокрылым, не обладают этой особенностью. В семье перепончатокрылых обычно бывает только одна половозрелая матка. Она совершает в молодом возрасте лишь один брачный вылет, запасаясь сперматозоидами на всю жизнь (на 10 лет и более). Эти сперматозоиды она использует постепенно на протяжении всех этих лет для оплодотворения яиц во время их прохождения по половым путям. Однако оплодотворяются не все яйца. Из неоплодотворенных яиц разбиваются самцы. Поэтому у самца нет отца и все его клетки тела, содержат лишь один набор хромосом (все они получены от матери) вместо двух (одного от матери и одного от отца), как, например, у человека. Прибегая к предложенной аналогии, можно сказать, что у самца перепончатокрылых в каждой клетке содержится лишь по одному экземпляру каждого «тома» вместо обычных двух.
Что касается самки перепончатокрылого, то она нормальна в том смысле, что у нее есть отец, и каждая клетка ее тела содержит обычный двойной набор хромосом. Превратится ли самка в рабочего или в матку, зависит не от ее генов, а от того, как ее выращивали. Это означает, что каждая самка несет полный набор генов, необходимый для того, чтобы стать маткой, и полный набор генов, необходимый для развития рабочего (или, вернее, наборы генов, детерминирующих каждую специализированную касту рабочих, солдат и т.п.). Какой набор генов будет реализован, зависит от условий, в которых выращивают самку, а в особенности пищи.
Такова в общих чертах система определения пола у перепончатокрылых, хотя имеется много осложняющих обстоятельств. Нам неизвестно, как возникла эта необыкновенная форма полового размножения. Несомненно, тому были веские причины, однако пока что нам следует относиться к ней как к любопытному факту биологии перепончатокрылых. Какими бы ни были первоначальные причины этого своеобразия, оно полностью разрушает изложенные четкие правила для вычисления коэффициента родства. Оно означает, что сперматозоиды отдельного самца вместо того, чтобы отличаться друг от друга, как у человека, совершенно одинаковы. В каждой клетке тела самца имеется лишь один, а не два набора генов. Поэтому каждый сперматозоид получает полный набор генов, а не 50%-ную выборку, и все сперматозоиды, продуцируемые данным самцом, идентичны. Постараемся теперь вычислить коэффициент родства между матерью и сыном. Если известно, что у данного самца имеется ген A , то какова вероятность наличия этого гена у его матери? Эта вероятность равна 100%, так как у самца не было отца, и все свои гены он получил от матери. Допустим теперь, что у некой матки имеется ген В . Вероятность того, что у ее сына будет этот ген, составляет всего 50%, поскольку сын содержит лишь половину генов матери. Это кажется противоречивым, но на самом деле противоречия здесь нет. Самец получает все свои гены от матери, но мать отдает сыну только половину своих генов. Разрешение этого кажущегося парадокса кроется в том, что клетки самца содержат лишь половину нормального числа генов. Нет смысла ломать себе голову над тем, чему равен «истинный» коэффициент родства — 1/2 или 1. Этот коэффициент — мера, придуманная человеком, и если в отдельных случаях он создает трудности, то следует отказаться от него и вернуться на исходные позиции. С точки зрения гена, находящегося в теле матки, вероятность наличия этого гена у одного из ее сыновей или дочерей равна 1/2. Поэтому с точки зрения матки ее потомки, независимо от их пола, находятся с ней в таком же близком родстве, как мать со своими детьми у человека.
Положение осложняется, когда дело доходит до сестер. Родные сестры не просто дочери одного и того же отца: зачавшие их сперматозоиды идентичны по всем своим генам. Поэтому сестры в том, что касается генов, полученных ими от родителей, равноценны однояйцовым близнецам. Если у одной самки имеется ген A , то она могла получить его либо от отца, либо от матери. Если она получила его от матери, то с вероятностью 50% он имеется и у ее сестры. Но если он достался ей от отца, то вероятность наличия этого гена у ее сестры равна 100%.
Поэтому у перепончатокрылых коэффициент родства между родными сестрами равен не 1/2, как у других животных с обычным половым размножением, а 3/4.
Отсю да сл едует, что у перепончатокрылых самка связана со своими сестрами более тесным родством, чем со своими потомками как одного, так и другого пола. По мнению Гамильтона (хотя он излагал его несколько иначе), вполне возможно, что это обстоятельство предрасполагает самку ухаживать за своей матерью как за эффективной машиной, производящей сестер. Ген, детерминирующий создание сестер косвенным образом, реплицируется быстрее, чем ген, детерминирующий непосредственное произведение на свет потомков. В результате в процессе эволюции возникла стерильность рабочих. Вероятно, нельзя считать случайностью, что истинно общественный образ жизни одновременно со стерильностью рабочих возникал у перепончатокрылых не менее чем 11 раз и притом всякий раз независимо, но лишь единственный раз во всем остальном животном мире, а именно у термитов.
Здесь имеется, однако, один подвох. Для того чтобы рабочие могли успешно ухаживать за своей матерью, всячески заботясь о ней как о машине, производящей сестер, они должны как-то сдерживать ее естественное стремление производить такое же число маленьких братьев. С точки зрения рабочего (напомним, что это самка) вероятность того, что каждый отдельный рабочий будет нести один определенный материнский ген, равна всего лишь 1/4. Поэтому, если бы матке была предоставлена возможность производить репродуктивных потомков в равных долях, то заботы о ней не окупались бы, во всяком случае, с точки зрения рабочих: все их труды не приводили бы к максимизации числа их драгоценных генов.
Трайверс и Хейр понимали, что рабочие, очевидно, стараются сдвинуть соотношение полов в пользу самок. Взяв расчеты Фишера для оптимального соотношения полов, они переработали их для особого случая перепончатокрылых. Оказалось, что стабильное отношение вклада для матери составляет, как обычно, 1:1. Однако стабильное соотношение для сестры равно 3:1 в пользу сестер, а не братьев. Для самки перепончатокрылых наиболее эффективный способ воспроизводства своих генов состоит в том, чтобы воздерживаться самой от размножения, предоставив своей матери производить для нее репродуктивных сестер и братьев в соотношении 3:1. Если, однако, самка должна иметь собственных потомков, то лучшее, что она может сделать для блага своих генов, это рожать репродуктивных сыновей и дочерей в равном соотношении.
Как уже было показано, различие между маткой и рабочими не является генетическим. В том, что касается генов, эмбрион самки может стать либо рабочей особью, которая «желает» соотношения полов 3:1, либо маткой, «желающей» соотношения 1:1. Что же означает это «желание»? Оно означает, что ген, находящийся в теле матки, воспроизводит себя наилучшим образом, если это тело вносит равные вклады в производство репродуктивных сыновей и дочерей. Но тот же самый ген, если он находится в теле рабочей самки, будет воспроизводиться лучше всего, если он заставит мать этого тела рожать больше дочерей, чем сыновей. В действительности здесь нет парадокса. Ген должен наилучшим образом использовать имеющиеся в его распоряжении рычаги власти. Если он может оказать влияние на развитие тела, которому суждено превратиться в матку, то он, чтобы использовать это влияние, прибегнет к одной оптимальной стратегии. Если же он в состоянии повлиять на способ развития тела рабочего, то он использует для этого другую стратегию.
Это означает, что на «ферме» перепончатокрылых существует столкновение интересов. Матка «старается» вносить равные вклады в самцов и самок. Рабочие пытаются сместить соотношение репродуктивных особей так, чтобы на каждого самца приходилось по три самки. Если мы вправе представить рабочих как фермеров, а матку — как племенную кобылу, то следует предполагать, что рабочие успешно достигнут желательного для них соотношения 3:1. Если же нет, если матка и в самом деле «царица», а рабочие — ее рабы и послушные няньки в царских яслях, то следует ожидать соотношения 1:1, которое «предпочитает» матка. Кто побеждает в этом особом случае битвы поколений? Данная ситуация поддается проверке, которую и предприняли Трайверс и Хейр, использовав большое число видов муравьев.
Среди соотношений полов интерес представляет соотношение репродуктивных самцов и самок. Периодически из гнезд муравьев происходят как бы выбросы крупных крылатых форм, отправляющихся в брачные полеты, после чего молодые матки предпринимают попытки основать новую семью. Эти-то крылатые формы и надо подсчитывать, чтобы оценить соотношение полов. У многих видов репродуктивные самцы и самки сильно различаются по размерам. Это осложняет дело, поскольку, как уже говорилось, расчеты Фишера относительно оптимального соотношения полов строго приложимы не к числу самцов и самок, а к количеству вкладов в самцов и самок . Трайверс и Хейр учли это, взвешивая муравьев. Они определяли соотношение полов по вкладам в репродуктивных индивидуумов у 20 видов муравьев. Их результаты достаточно близки к соотношению 3 самки: 1 самец, предсказанному теорией, т.е. подтверждают, что рабочие муравьи командуют парадом в собственных интересах.
Создается, таким образом, впечатление, что у изученных муравьев в конфликте интересов «побеждают» рабочие. Это не слишком удивляет, поскольку тела рабочих, охраняющих питомники, в практическом отношении обладают большей силой, чем тела маток. Гены, пытающиеся подчинить себе мир через тела маток, не могут одолеть гены, властвующие над миром через тела рабочих. Интересно было бы найти какие-то особые обстоятельства, при которых матки могли бы обладать большей практической властью, чем рабочие. Трайверс и Хейр поняли, что существует одно именно такое обстоятельство, которое можно использовать для критической проверки теории.
Оно связано с тем, что некоторые виды муравьев заводят себе рабов. У таких видов рабочие либо вовсе не занимаются своим обычным делом, либо выполняют его плохо. Зато они очень хорошо умеют захватывать рабов. Настоящие войны, в которых большие армии противников дерутся насмерть, известны только у человека и у общественных насекомых. У многих видов муравьев специализированная каста, известная под названием солдат, вооружена устрашающими челюстями и отдает все свое время сражениям за свою семью против армий других муравьев. Набеги за рабами — это просто одно из военных действий. Охотники за рабами нападают на гнездо муравьев другого вида, убивают рабочих или солдат, которые его защищают, и уносят с собой расплод. Молодь вылупляется в гнезде своих поработителей и, не «понимая», что попала в рабство, начинает действовать в соответствии со встроенными в ее нервную систему программами. Рабочие, захватывающие рабов, или солдаты продолжают совершать набеги за новыми рабами, а в их гнездах рабы выполняют повседневные функции, связанные с уборкой и уходом за молодью.
Рабы, конечно, находятся в счастливом неведении о том, что они не связаны родством ни с матерью, ни с молодью, за которой они ухаживают. Невольно они выращивают новые отряды рабовладельцев. Не может быть сомнений, что естественный отбор, действуя на гены рабского вида, благоприятствует «антирабским» адаптациям. Однако эти адаптации, по-видимому, недостаточно эффективны, потому что рабство оказалось широко распространенным явлением.
С рассматриваемых здесь позиций интерес представляет следующее последствие рабства: матка вида, захватывающего рабов, получает возможность сдвигать соотношение полов в ту сторону, которую она «предпочитает». Это связано с тем, что ее собственные, действительно рожденные ею потомки-захватчики рабов уже не имеют практической власти в яслях. Власть эта оказалась в руках рабов. Рабы «думают», что они ухаживают за своими собственными личинками и, надо полагать, делают все то, что им надлежало бы делать в своих собственных гнездах для достижения желаемого соотношения 3:1 — в пользу сестер. Но матка рабовладельческого вида в состоянии принять контрмеры, нейтрализовать которые рабы не могут, так как между ними и молодью нет никакого родства и они соответственно не подвергаются отбору.
Допустим, например, что у какого-то вида муравьев матки «пытаются» замаскировать яйца мужского пола, придавая им запах яиц женского пола. В нормальной ситуации естественный отбор поддержит любую тенденцию рабочих «разгадать» обман. Можно представить себе эволюционную битву, в которой матки постоянно «меняют код», а рабочие «расшифровывают» его. Выиграет битву тот, кто сумеет передать следующему поколению больше своих генов через тела репродуктивных индивидуумов. Как мы убедились, побеждают обычно рабочие. Но когда код изменяет матка рабовладельческого вида , то среди рабочих-рабов не может произойти отбор на способность к его расшифровке, потому что ген, детерминирующий эту способность, не содержится ни в одном из репродуктивных индивидуумов, а, следовательно, не может передаваться по наследству. Все репродуктивные индивидуумы принадлежат к рабовладельческому виду, т.е. связаны родством с маткой, но не с рабами. Если гены рабов и попадают в каких-то репродуктивных индивидуумов, то последние непременно должны происходить из того гнезда, из которого были похищены сами эти рабы. Так что рабочие-рабы в лучшем случае будут заниматься расшифровкой кода, не имеющего к ним никакого отношения! Поэтому матки рабовладельческого вида могут безнаказанно изменять свои гены, отнюдь не рискуя, что гены, детерминирующие способность к расшифровке кода, будут переданы следующему поколению.
Результат этих сложных рассуждений сводится к тому, что у рабовладельческих видов соотношение вкладов в репродуктивных индивидуумов двух полов, очевидно, должно быть ближе к 1:1, а не к 3:1. Хоть в одном случае матка сможет поступать так, как ей угодно. Именно это и установили Трайверс и Хэйр, хотя они изучали только два рабовладельческих вида.
В реальной жизни не все так просто и ясно. Например, самый хорошо знакомый нам вид общественных насекомых — медоносная пчела — ведет себя, казалось бы, совсем «неправильно». Вклад в трутней значительно превышает у нее вклад в маток, что представляется нелепым с точки зрения, как рабочих пчел, так и самой матки. Гамильтон предложил возможное решение этой загадки. Он указал, что когда пчелиная матка покидает улей, ее сопровождает большая толпа рабочих, которые помогают ей основать новую семью. Для родного улья эти рабочие потеряны навсегда и затраченные на них ресурсы следует включить в стоимость размножения: на каждую матку, покидающую улей, необходимо произвести на свет большое число дополнительных рабочих. Вклады в этих дополнительных рабочих пчел следует рассматривать как часть вклада в репродуктивных самок. При вычислении соотношения полов дополнительных рабочих пчел следует приплюсовать к маткам. Так что, в конечном счете, это не такое уж серьезное затруднение для теории.
Большая проблема для этой элегантной теории возникает в связи с тем, что у некоторых видов молодая матка во время своего брачного полета спаривается не с одним, а с несколькими самцами. Это означает, что коэффициент родства между ее дочерьми в среднем ниже, чем 3/4, а в некоторых экстремальных случаях может даже приближаться к 1/4. Соблазнительно, хотя, возможно, не очень логично, рассматривать это как хитрый удар, наносимый матками рабочим! Можно было бы думать, что рабочие пчелы должны сопровождать матку в ее брачном полете, чтобы помешать ей спариваться больше одного раза. Но это никак не поможет собственным генам рабочих — только генам следующего их поколения. Среди рабочих как класса нет духа профессиональной солидарности. Каждый из них заботится лишь о собственных генах. Рабочей пчеле, быть может, «хотелось бы» сопровождать свою собственную мать, но ей не представилось такой возможности, поскольку она не была зачата в эти дни. Молодая матка, совершающая свой брачный полет, приходится имеющемуся на данный момент поколению рабочих пчел сестрой, а не матерью. Поэтому они будут на ее стороне, а не на стороне следующего поколения рабочих, которые приходятся им всего лишь племянницами.
ШМЕЛЬ: МНОГО ШУМА НИ ИЗ ЧЕГО
Шмели относятся к перепончатокрылым, как и пчелы, с которыми они находятся в близком родстве и имеют во многих отношениях похожие нравы и повадки. И у пчел, и у шмелей рождаются самцы, не имеющие жала, но у пчелиных самцов, трутней, нет тяги к путешествиям, они не отлетают далеко от улья, где собраны громадные запасы корма, которым они и питаются до осени, когда безжалостные самки выгоняют их прочь из улья, обрекая на смерть от холода и голода.
У шмелей все иначе. В их гнезде никогда не бывает слишком много меда. Они не делают зимних запасов, разве что на несколько дней или недель - на случай плохой погоды, когда, как известно, шмели не вылетают на поиски корма.
Поедать запасы из ячеек, в которых кормится молодежь? Нет, этого не допустили бы шмели-няни, поэтому самцы тут появляются поздно, только где-то в августе. Кроме того, они гораздо более активные и самостоятельные, чем пчелиные трутни. Они летают далеко от гнезда и берут взяток в виде пыльцы и нектара, но только для сиюминутного поедания на месте - ни один из них, как и пчелиные трутни, никогда не принес ни единого зернышка в общественную кладовую.
Таким образом, небольшие запасы, имеющиеся в шмелином гнезде, служат только для кормления детей, на зиму они не пригодятся, поскольку уже поздней осенью все семейство гибнет: и старая матка, и трутни, и работницы.
Кто же останется?
Остаются только оплодотворенные в августе самки, которые, как многие другие насекомые, находят себе теплое убежище и проводят в нем зиму, погруженные в оцепенение до первых теплых солнечных лучей.
И вот тогда шмелиной королеве приходится немало потрудиться. Она одна на свете, и на ее плечи ложится и закладка роя, и выкармливание первых его работниц.
Прежде всего, она находит какую-нибудь ямку в земле, брошенную норку крота или мыши, щель между камнями, птичье гнездо или скворечник и начинает там выкладывать пол из воска, вырабатываемого железами на верхней и нижней стороне ее брюшка и смешиваемого с пыльцой и даже с измельченными кусочками растений.
На этом основании она строит круглую стенку, уже из чистого воска, заполняет ее внутри пыльцой и откладывает туда несколько яиц, а затем замуровывает эту ячейку. Рядом с первой она строит вторую, третью и т.д.
Но в отличие от некоторых пчел одиночек, которые, заперев своих детей в камере с едой, больше ими не интересуются, шмелиная королева открывает время от времени первые ячейки и проверяет, хватает ли корма личинкам, пополняя запас в случае необходимости, и так до момента, когда личинки превращаются в куколок, сидящих в бронзовых коконах.
И тут наступает интересный момент: матка энергично разбивает стенку ячейки, чтобы использовать этот воск для строительства новых ячеек. Чтобы производить новый строительный материал, она должна хорошо питаться, а у нее просто нет на это времени в разгар строительных работ.
Кто-то появился на свет
Наконец из коконов вылезают первые помощницы, очень странные на вид. Безусловно, это шмели, шмели самки, но они втрое меньше матери, какие-то невзрачные карлицы. У них также есть жало, но слишком маленькое, чтобы причинить боль человеку.
Как бы то ни было, наконец-то появляется какая-то помощь для заваленной работой королевы. Малыши строят, носят корм, а мать хотя и работает вместе с ними, но может уже больше времени посвящать кладке яиц. Число маленьких помощниц все растет и к тому же увеличиваются их размеры, ибо корма стало больше.
Рой день ото дня становится тяжелее. Матка наконец-то может освободиться от личного участка в добывании корма. Этим занимаются исключительно ее донки, а она целиком отдается кладке яиц, которых производит все больше и больше.
Хорошо откормленные личинки начинают, в свою очередь, вырастать до размеров матки роя. Они тоже будут королевами, потому что уже приближается осень, и среди потомства начинают появляться самцы. Как и у пчел, для рождения трутня не требуется участия самца, так как появляются они из неоплодотворенных яиц, которые могут в то же лето отложить также самые большие из вновь родившихся самочек.
Приходит время брачных полетов, после которых самцы попросту погибают, а оплодотворенные самочки начинают подыскивать места для зимовки и, как правило, уже не возвращаются в родное гнездо.
А там дела идут все хуже. Самки работницы приносят все меньше провизии, так как цветов в полях становится меньше, и в результате все семейство - и старая королева, и работницы разных размеров, которые не принимали участия в брачных полетах из-за недоразвитости их органов размножения, и конечно, все самцы - погибают от голода и холода.
На другой год вся история начинается сначала, причем все заботы по созданию роя снова ложатся "на плечи" несчастной шмелиной королевы. Зато пчелиная королева-мать не будет участвовать весной в поисках корма для себя и своих будущих детей, поскольку в улье с богатыми запасами еды перезимовал вместе с ней целый рой ее работящих дочек, которые с первых теплых дней начинают полеты за провиантом.
Если прибавить к этому, что ячейки шмелей имеют неуклюжий, бесформенный вид, что сосуды для меда похожи ни уродливые бочонки, и в качестве посуды используются иногда остатки коконов от куколок, то становится видно, как далеко шагнули вперед пчелы в отношении бытового благоустройства, а значит, и в борьбе за жизнь, по сравнению со своими мохнатыми родичами.
У шмелей есть свои любимые растения. Ранней весной они летают над вербами, под осень особенно охотно кормятся с чертополоха. Здесь нельзя не отметить большое значение шмелей для сельского хозяйства, поскольку, располагая гораздо более длинным язычком, чем пчелы, они могут забираться за взятком в такие глубокие цветочные чашечки, которые дня пчел недосягаемы. Например, пчелы не могут справиться с красным клевером и полому его избегают, так что иногда приходится специально дрессировать пчел-работниц в этом направлении.
Шмель же забирается в этот цветок без всякой дрессировки, часто помогая себе в этом таким образом: снабженный довольно сильными челюстями, он просто выгрызет сбоку дырочку, чтобы добраться до нектарника. В любом случае он способствует опылению клевера.
Шмелиные куколки
Любопытно, что при своей не слишком высоко организованной общественной жизни шмели уже обзавелись в своей семье паразитическими видами. Таким социальным паразитом является шмель из рода Psithyrus, очень похожий на нашего обычного земляного шмеля с той разницей, что не производит работниц, а только самцов и самок.
Самцы, как обычно, интересуются только брачным полетом и живут только до времени этого полета. Самки же, обычно такие трудолюбивые, у этого вида ведут себя совсем иначе. Они проникают в ячейки, приготовленные для потомства нормальной шмелиной семьи, и откладывают в них свои яйца, совсем как кукушки. И земляные шмели выкармливают чужаков, как птицы кукушат.
Шмели описанных видов живут не только в Европе, но и в Азии, и в Америке, забираясь иногда так далеко на север, что им не хватает времени на весь цикл воспроизводства работниц, и тогда шмелиная матка ведет себя, как пчела-одиночка, а именно - она откладывает только одну партию яиц и оставляет их на произвол судьбы.
Зато на юге случается иногда, что при условии теплых зим шмели ведут свой нормальный образ жизни круглый год.
Как видим, эти жужжащие толстяки умеют довольно хорошо приспосабливаются к климату.
По книге Яна Жабиньски
"Из жизни животных"
Укус шершня убил рыбака
Трагически закончилась для жителя Нововятского района поездка на рыбалку в Уржум. В жаркий солнечный день укус шершня оказался для него смертельным. Пострадавший был защищен рубашкой и брюками, и все же насекомое умудрилось забраться именно под одежду. Последовала аллергическая реакция организма, аритмия сердца - и через пять минут человека не стало.
Шершни - род жалящих насекомых из отряда перепончатокрылых длиной до 40 мм. Укус болезнен. Питаются насекомыми, в том числе пчёлами.
Аллергологи данный случай для нашей местности считают из ряда вон выходящими. Летальные исходы до сих пор фиксировались только после нападения пчёл.
Страдающим подобного рода аллергическими реакциями, а также людям, неоднократно укушенным осами или пчёлами, специалисты рекомендуют иметь при себе медицинскую аптечку. В её содержимое входят шприцы для инъекций, адреналин, преднизолон, димедрол, супрастин и другие препараты.
Выписка из газеты "Вятский Наблюдатель", N 32, 6 августа 1999 г.